-
HOME
Educação sem fronteiras
-
ALUNOS
A faculdade ao seu alcance

Assine o Educabras!
Próxima parada, faculdade
Nosso objetivo é prepará-lo para o Vestibular e o Enem, para que você possa entrar na faculdade de sua escolha e seguir a carreira de seus sonhos.
Torne-se um assinante a partir de R$29,90/mês Mais Informações

-
PROFESSORES
agilize seu trabalho
Assine o Educabras!
Solução inteligente
Fácil de usar, eficaz e econômico, o Educabras.com é uma solução para professores de Ensino Médio, Pré-Vestibular e Enem.
-
COLÉGIOS
Planos diferenciados

Para colégios, cursos e ONGs.
Colégio conectado
Duas opções:
1. Assinaturas a preços com desconto.
2. Uma única assinatura que inclui o gerador de provas de todas as matérias. -
Área Livre
Aulas e Blog

Área livre do Educabras.com, aberta para assinantes e visitantes.
Educação é essencial
Educação, Conhecimento e diversos outros assuntos relevantes para alunos e professores.
-
Entre em contato
Tire suas dúvidas


Processo da Fotossíntese
- Home
- Processo Da Fotossíntese
Fotossíntese
Plastos
Os plastos são estruturas exclusivas de algas e vegetais. O seu número e forma varia muito conforme o organismo estudado.
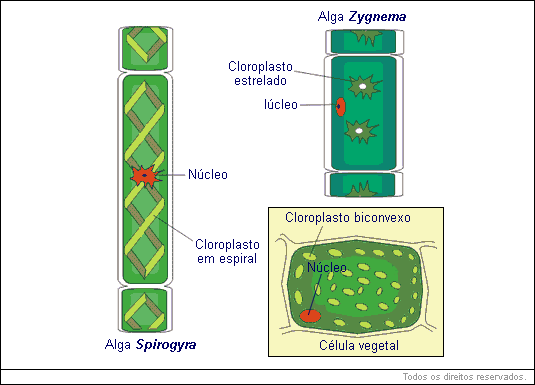
Existem basicamente dois tipos de plastos: cromoplastos e leucoplastos. Os cromoplastos apresentam pigmentos no seu interior (cromo = cor), os leucoplastos (leuco = branco), não contêm pigmentos. O cromoplasto mais comum nos vegetais é o cloroplasto.
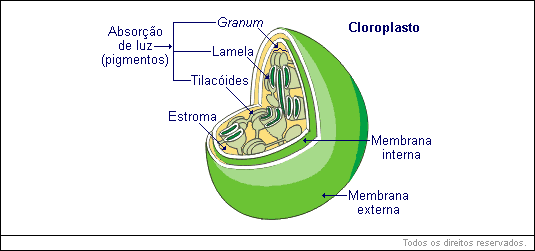
Cloroplastos
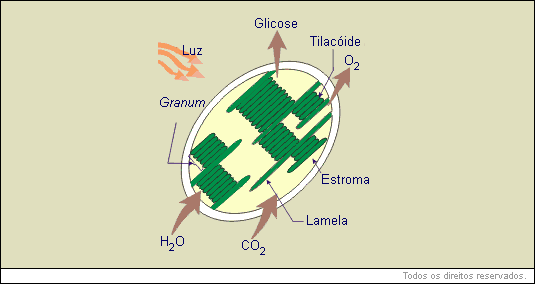
Os cloroplastos apresentam forma discoidal, são envolvidos por uma membrana externa e uma interna. Além destas, os plastos apresentam muitas membranas internas que formam bolsas chatas em forma de disco chamadas tilacoides. Estes, por sua vez, estão dispostos de modo a formar pilhas, semelhantes a uma pilha de moedas. A pilha de tilacoides recebe o nome de granum (plural = grana) . O interior do cloroplasto é preenchido por uma matriz gelatinosa chamada estroma, onde se encontram DNA, RNA, ribossomos, enzimas, etc.
Nos tilacoides estão localizadas as moléculas de clorofilas, as quais estão organizadas de modo a poderem captar a maior quantidade de luz possível.
Funções
Nos cloroplastos acontece a fotossíntese, processo onde são fabricadas moléculas orgânicas, principalmente glicose, usada pelas mitocôndrias na respiração intracelular.
Durante a fotossíntese a clorofila capta a energia luminosa que será transformada em energia química (ATP). Essa energia será usada na fabricação de glicose a partir de água e gás carbônico.
Cloroplastos ou leucoplastos podem armazenar o excesso de glicose produzida em forma de amido (polissacarídeo). Esses reservatórios são os amiloplastos.
Como as mitocôndrias, os cloroplastos são capazes de se autoduplicar independentemente da duplicação celular e sintetizar alguns tipos de proteínas.
Fotossíntese
Os vegetais clorofilados têm o equipamento bioquímico necessário para transformar substâncias pouco energéticas (CO2 e H2O) em substância rica em energia (glicose). Na fotossíntese, a energia luminosa absorvida pela clorofila é transformada em energia química de ligação, que fica armazenada no carboidrato.
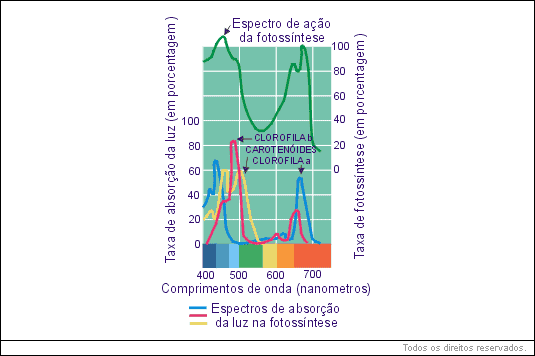
A luz utilizada nessa formação é absorvida por uma série de pigmentos. Cada pigmento absorve determinados comprimentos de ondas, refletindo os que não absorve. A cor do pigmento é dada pelo comprimento de onda refletido, podendo-se determinar o espectro de absorção de cada pigmento através de um espectrofotômetro.
Os tipos de pigmentos utilizados na fotossíntese variam nos diferentes grupos de organismos fotossintetizantes. Nos vegetais superiores, os pigmentos mais importantes são a clorofila a e a clorofila b, pigmentos verdes que absorvem a luz no violeta, no azul e no vermelho, refletindo no verde; por isso, são verdes.
Colocando-se em um gráfico os diferentes comprimentos de onda em função da taxa em que se processa a fotossíntese, pode-se verificar o espectro de ação da luz na fotossíntese:
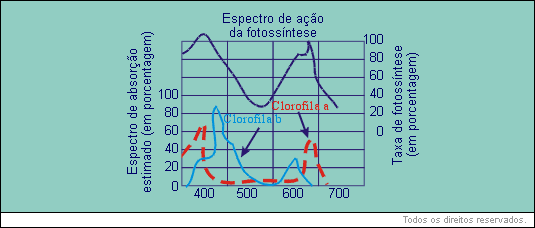
Observando-se os gráficos apresentados, pode-se notar que os picos do espectro de ação da luz na fotossíntese e os dos espectros de absorção da luz pela clorofila têm padrão semelhante, evidenciando que a clorofila é o pigmento mais importante na recepção da luz na fotossíntese.
A absorção da luz pela clorofila se faz com intensidade máxima nas faixas de comprimento de onda de 450 nm (nanômetros), que é correspondente à luz azul, e 700 nm que corresponde, à luz vermelha. O nanômetro ainda é muito conhecido como milimícron. A absorção da luz verde é quase nula. A clorofila reflete-a quase integralmente. E é por isso que nós a vemos dessa cor.
As curvas indicam:
a) espectro de absorção da luz pela clorofila a;
b) espectro de absorção da luz pela clorofila b;
O espectro de ação da fotossíntese não corresponde rigorosamente ao espectro de absorção da luz pelas clorofilas. Considerando o fenômeno fotossíntese em termos gerais, a resposta à luz vermelha é maior do que à luz azul. Isso se explica porque os pigmentos acessórios (xantofila, licopeno e caroteno) absorvem intensamente certas radiações de forma mais eficiente que as clorofilas, transferindo depois a elas a energia absorvida.
A equação tradicional da fotossíntese é:
![]()
Essa reação, no entanto, não pode mais ser aceita como correta, tendo em vista que o oxigênio liberado na fotossíntese provém da água e não do gás carbônico. Isto foi confirmado por um experimento clássico (década de 40), no qual o oxigênio da água foi marcado com o isótopo O18, verificando-se que todo o oxigênio liberado na fotossíntese era isótopo18.
Dessa forma, a reação aceita é:
![]()
Essa equação mostra o processo de síntese de compostos orgânicos a partir de substâncias inorgânicas, utilizando-se a energia luminosa e com liberação de oxigênio (O2).
Hoje, sabemos que a fotossíntese se processa em duas etapas. Na primeira, a luz cede energia para a clorofila. Portanto, essa etapa não ocorre sem a presença de luz. Ela é conhecida com fase luminosa da fotossíntese ou reações de claro.
Nessa fase ocorre a fotólise de Hill ou fase fotoquímica. Nela é liberado o O2 da água, e os hidrogênios são incorporadas a um aceptor de hidrogênio, no caso, o NADP (nicotinamida - adenina - dinucleotídeo - fosfato).
A reação da primeira etapa é:

Além disso, ocorre a formação de ATP pela utilização direta da energia luminosa - fotofosforilação.
Na segunda etapa, a energia retida por certos compostos, vai permitir uma série de reações que vão levar ao aparecimento da glicose. Essa etapa pode ocorrer mesmo na ausência da luz. É a fase escura ou reações de escuro da fotossíntese. A segunda etapa é chamada puramente química, proposta por Blackman, na qual o NADPH2, reduz o CO2, formando-se açúcar.
Essa reação é:
![]()
Somando-se as duas reações:

Durante a fase obscura, o NADPH2 e o ATP produzidos durante as reações fotoquímicas são utilizados para reduzir o CO2. Ocorrem no estroma do cloroplasto. Foram os trabalhos de Calvin, Bassham e Benson, empreendidos desde 1946, que permitiram conhecer as diversas etapas da redução do gás carbônico a glicídios.
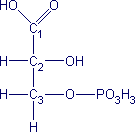
Com métodos, combinando a autorradiografia e a cromatografia, Calvin e seus alunos mostraram que o primeiro composto estável que aparece, após somente alguns segundos de fotossíntese, em presença de CO2 marcado, é uma substância em C3 o ácido fosfoglicérico (PGA) do qual um dos carbonos, o do grupamento carboxila é radioativo.
A fórmula do PGA é a seguinte:
Contrariamente ao que se pensava, o CO2 não se fixa numa substância em C2 para dar o PGA, mas num açúcar em C5 fosforilado: a ribulose - 1,5 - difosfato (RuDP). Fixando-se na ribulose, o CO2 dá origem a um composto de seis carbonos muito instável, que se decompõe imediatamente em duas moléculas de ácido fosfoglicérico (PGA), conforme a reação seguinte:
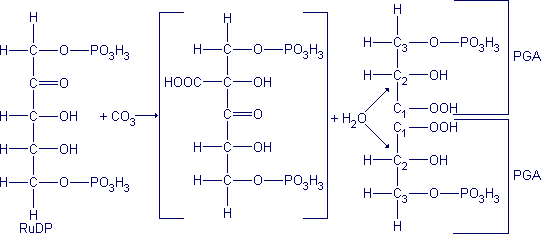
A partir de duas moléculas de ácido fosfoglicérico (PGA) forma-se glicose, por uma série de reações cujo desenrolar é o inverso da glicólise. É durante essa fase que são utilizados uma parte do NADPH2 e uma parte do ATP, formados na fase luminosa.
A oxidação do NADPH2 em NADP e a hidrólise do ATP em ADP mais P inorgânico permitem a formação de triose fosfato. Não só açúcares se formam a partir do PGA, mas também aminoácidos, ácidos orgânicos, ácidos graxos e glicerol.
Os fotossistemas I e II
Essa descrição servirá como "enriquecimento" para visualização da alta complexidade com que o processo biológico fotossintético se desenvolve. Procure conhecê-lo em linhas gerais!
Modernamente tem se falado em fotossistemas, para designar unidades funcionais presentes nas membranas dos tilacoides constituídas por moléculas de pigmentos, associadas entre si. Nesses fotossistemas, há dois tipos de clorofilas (a e b) e pigmentos amarelos e laranjas, os carotenoides. São conhecidos dois tipos de fotossistemas, I e II que, absorvem comprimentos de onda luminosa ligeiramente diferentes e agem juntos na fotossíntese. Em ambos, a clorofila a parece ser a molécula fundamental.
A fotólise da água está associada à fotofosforilação acíclica.
Sabe-se hoje que no cloroplasto, além da produção cíclica de ATP, que já conhecemos, há produção de ATP acoplada à "quebra" da água . O processo é razoavelmente complexo:
1.O fotossistema II (clorofila b) recebe luz. Elétrons ganham energia e são capturados por aceptor. A água perde elétrons, que são cedidos ao fotossistema II, e origina íons H+ que ficam no meio e O2 liberado da planta.
2. Os elétrons que saíram do fotossistema II são transferidos a uma cadeia de aceptores, alguns dos quais são citocromos. Na sua passagem "ladeira abaixo" eles perdem energia, usada na produção de ATP.
3. O fotossistema I (clorofila a) também foi iluminado e perdeu seus próprios elétrons para uma molécula aceptora. Por essa razão, ele se torna aceptor e recebe os elétrons provenientes do fotossistema II.
4.Os elétrons provenientes do fotossistema I, junto com íons H+ provenientes da água, são cedidos ao NADP, que se transforma em NADPH2.
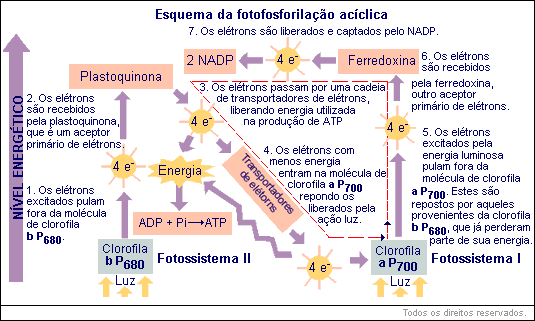
Você então percebe que, juntamente com a fotólise da água, ocorreu produção de ATP. Dizemos que esta produção é acíclica, porque os elétrons que a permitiram provieram da água e terminaram nos NADPH2.
A fosforilação cíclica também pode ser enxergada nesse esquema. Ela é representada pela linha pontilhada. Em alguns casos, os elétrons do fotossistema I, ao invés de seguirem para o NADP, são entregues à cadeia de transportadores e voltam ao próprio fotossistema I. Tanto o ATP feito ciclicamente como aquele que é produzido simultaneamente à quebra da água serão usados na fase de "escuro".
Resumindo:
|
O fotossistema II, estimulado pela luz, solta elétrons ricos em energia que passam por aceptores e permitem produção acíclica de ATP. O fotossistema II é regenerado ganhando elétrons da água. O fotossistema I, quando iluminado, cede elétrons de alta energia ao NADP que, junto com íons 2H+ da água, se transforma em NADPH2. Os elétrons do fotossistema II regeneram o fotossistema I. Um caminho alternativo dos elétrons entre o fotossistema I e os aceptores gera produção (cíclica) de ATP. |
Visualização geral
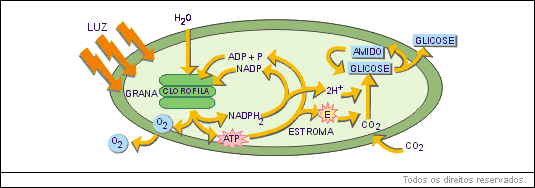
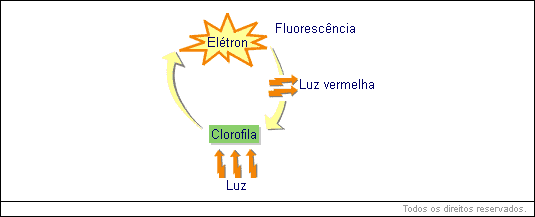
Poderíamos, então, nos perguntar se o cloroplasto íntegro contém clorofila, por que motivo não fluoresce (emite luz vermelha), quando iluminado?
A análise química do cloroplasto demonstrou apresença de várias substâncias aceptoras de elétrons, entre as quais alguns citocromos. Determinou-se então que, no cloroplasto iluminado, deve ocorrer algo semelhante ao esquema abaixo.
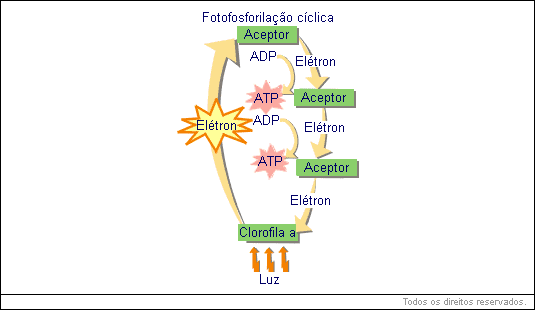
Note que também aqui o elétron da clorofila a iluminada ganha energia; porém este elétron é removido da clorofila pelo primeiro receptor. Em seguida, o elétron "desce" a níveis energéticos menores, sendo capturado por aceptores intermediários ("degraus energéticos") antes de retornar à clorofila. Nesta "descida", a energia perdida pelo elétron é capturada, resultando na produção de ATP.
Compare agora os esquemas anteriores da fluorescência e da fotofosforilização: na fluorescência, já que o elétron voltou diretamente à clorofila, toda energia absorvida por ele foi devolvida de uma vez só, sob a forma de luz (vermelha). Na fotofosforilização, o elétron perdido também acaba voltando à clorofila; porém não diretamente, e sim passando por substâncias intermediárias.
Neste caso, a energia do elétron é "perdida" aos poucos, o que permite sua captura pelo sistema ADP - ATP.
Você notou que o caminho do elétron é cíclico, já que ele sai da clorofila e acaba voltando a ela. Por este motivo, o processo é chamado fotofosforilação cíclica.
Não percamos de vista que o ATP produzido na fase de claro será utilizado na produção de glicose, na fase de escuro.
Resumindo:
|
Sabe-se hoje que o cloroplasto iluminado é capaz de fabricar ATP. Nele, a clorofila a excitada pela luz, perde um de seus elétrons, que ganha energia, sai da molécula, e é capturado por um aceptor. O elétron passa por aceptores intermediários e acaba voltando à clorofila, perdendo aos poucos a energia adquirida. Esta energia é utilizada na produção de ATP. Já que o caminho do elétron é "fechado", o processo é chamado fotofosforilação cíclica. |
Sumário
- Plastos- Cloroplastos
- Fotossíntese
i. Os fotossistemas I e II
Áreas exclusivas para assinantes