-
HOME
Educação sem fronteiras
-
ALUNOS
A faculdade ao seu alcance

Assine o Educabras!
Próxima parada, faculdade
Nosso objetivo é prepará-lo para o Vestibular e o Enem, para que você possa entrar na faculdade de sua escolha e seguir a carreira de seus sonhos.
Torne-se um assinante a partir de R$29,90/mês Mais Informações

-
PROFESSORES
agilize seu trabalho
Assine o Educabras!
Solução inteligente
Fácil de usar, eficaz e econômico, o Educabras.com é uma solução para professores de Ensino Médio, Pré-Vestibular e Enem.
-
COLÉGIOS
Planos diferenciados

Para colégios, cursos e ONGs.
Colégio conectado
Duas opções:
1. Assinaturas a preços com desconto.
2. Uma única assinatura que inclui o gerador de provas de todas as matérias. -
Área Livre
Aulas e Blog

Área livre do Educabras.com, aberta para assinantes e visitantes.
Educação é essencial
Educação, Conhecimento e diversos outros assuntos relevantes para alunos e professores.
-
Entre em contato
Tire suas dúvidas


Mapas genéticos
- Home
- Mapas Genéticos
Linkage (ligação ou vinculação)
A segunda lei de Mendel é válida quando estudamos dois ou mais pares de genes que estão situados em cromossomos homólogos diferentes.
Porém, a segunda lei de Mendel não se aplica a pares de genes situados no mesmo par de cromossomos homólogos, pois esses genes não terão segregação independente. Genes presentes no mesmo cromossomo tendem a permanecer juntos durante a formação dos gametas.
Essa explicação foi dada por Morgan, em 1910, e é chamado linkage ou vinculação. Morgan supôs que a tendência de os genes vinculados conservarem suas combinações originais era devida a se encontrarem ligados no mesmo cromossomo.
Além disso, emitiu a ideia básica de que o grau ou força de ligação depende da distância que separa os genes vinculados no cromossomo. A partir disso evoluiu a teoria da disposição linear dos genes nos cromossomos e depois partiu-se para a elaboração dos mapas genéticos ou dos genes ligados aos cromossomos.
A frequência de recombinação de um par de genes ligados é, em condições uniformes do ambiente, constante e característica de cada par de genes. Morgan emitiu a hipótese (1911) de que a intensidade do linkage de um par de genes – isto é, a frequência de recombinações observadas – depende da distância entre estes genes no cromossomo.
Os genes muito próximos um do outro, no mesmo cromossomo, estarão fortemente ligados e não apresentarão recombinação; isso é chamado de ligação completa. Entretanto, genes ligados nem sempre ficam juntos, devido ao fato que as cromátides homólogas poderem trocar pedaços entre si (fenômeno conhecido como crossing-over ou permutação) durante a prófase I da meiose. Quanto maior for essa distância, mais provável será, em geral, que ocorra recombinação entre os dois genes; a isso chamamos de ligação incompleta.
Ligação no milho
O fenômeno do linkage é particularmente claro no milho, no qual a recombinação ou permutação entre os genes ligados ocorre em ambos os sexos e onde os caracteres observáveis nas sementes contam-se em grande número, já que cada espiga forma numerosas sementes.
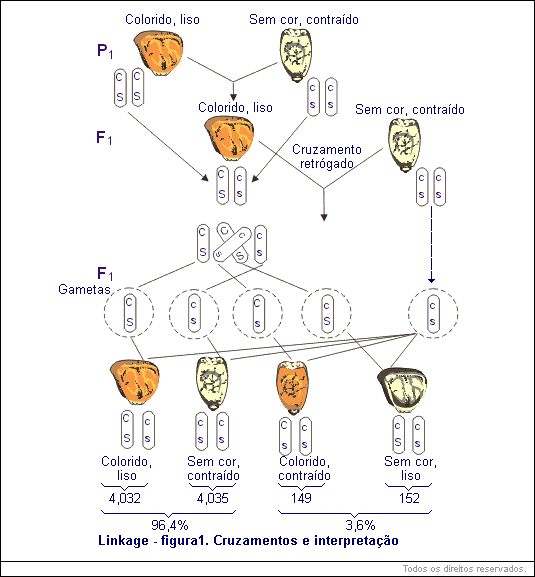
Constituem um ótimo exemplo os resultados obtidos por Hutchinson, que cruzou uma variedade de milho com sementes coloridas e normalmente cheias (lisas) com outra variedade incolor e de sementes contraídas (menos endosperma).
Em outros experimentos havia sido demonstrado que o gene C para cor, era dominante sobre c, falta de cor, enquanto o gene S, endosperma normal (liso) ou cheio, é dominante sobre contraído, s. De acordo com isso, os parentais CC SS (sementes coloridas e cheias) e cc ss (sementes incolores e contraídas), e a geração F1, como era esperado, apresentavam as sementes coloridas e cheias e o genótipo CS / cs:
|
Para representar os genes que se encontram em linkage, as combinações gênicas (pares de alelos) são escritas como acontecem no zigoto (Cc Ss), porém as de um parental sobre uma linha horizontal (CS ) e as do outro parental (cs), debaixo dela:
|
Se C e S fossem transmitidos independentemente, de acordo com a 2a lei de Mendel, estas plantas F1 (Cc Ss) deveriam produzir quatro tipos de gametas CS, Cs, cS e cs em igual proporção (¼ ou 25% de cada).
A maneira mais simples de comprovar essa proporção gamética é fazer um cruzamento-teste do resultado F1 com o duplo recessivo cc ss, o qual, por esta previsão indicada, produziria 4 classes de descendentes na proporção 1 : 1 : 1 : 1, ou seja, ¼ ou 25% de cada.
Ao se fazer esse cruzamento-teste, no entanto, não foi cumprida esta previsão, e sim foram obtidos os seguintes resultados:
Tabela 1
|
classes fenotípicas |
genótipos |
no de sementes |
proporção (%) |
|
coloridas e lisas |
CS / cs |
4.032 |
4032 / 8368=48,18% |
|
coloridas e contraídas |
Cs / cs |
149 |
149 / 8368= 1,78% |
|
incolores e lisas |
cS / cs |
152 |
152 / 8368= 1,81% |
|
incolores e contraídas |
cs / cs |
4.035 |
4035 / 8368=48,21% |
|
Total |
8.368 |
As sementes coloridas e lisas e as sementes incolores e contraídas apresentam-se em proporção maior que as sementes incolores e contraídas e incolores e lisas (conforme a tabela anterior). Lembremos que os parentais tinham as sementes coloridas e lisas (Cc Ss) e incolores e contraídas (cc ss). Portanto, estas combinações de caracteres são as parentais, enquanto que nas recombinações as associações de caracteres se apresentaram permutadas (coloridas e contraídas; incolores e lisas).
Se a transmissão fosse de caracteres independentes, as combinações parentais CS e cs com as recombinações Cs e cS, apareceriam com a mesma frequência (25% de cada). Contudo, no experimento indicado, a situação foi a seguinte:
Combinações parentais: 
Recombinantes: 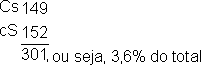
Fica evidente que nos parentais, os pares de genes C-c e S-s não foram transmitidos independentemente. As combinações parentais excederam em muito os 50% previstos, permanecendo ligadas ou vinculadas em 96,4% e recombinadas (permutadas) em somente 3,6% dos gametas.
Ao preparar outro experimento com os mesmos genes, porém com combinações iniciais diferentes, isto é, ao cruzar parentais de sementes incolores e lisas com outros de sementes coloridas e contraídas, foi visto que novamente as combinações parentais se encontravam em maior proporção (acima de 50%), embora fossem opostas às obtidas no primeiro experimento.
Os resultados do segundo experimento foram os seguintes:
Tabela 2
|
classes fenotípicas |
genótipos |
no de sementes |
proporção (%) |
|
coloridas e lisas |
CS / cc |
639 |
639 / 44595=1,43% |
|
coloridas e contraídas |
Cs / cs |
21.379 |
21379 / 44595=47,94% |
|
incolores e lisas |
cS / cs |
21.906 |
21906 / 44595=49,12% |
|
incolores e contraídas |
cs / cs |
672 |
672 / 44595= 1,51% |
|
Total |
8.368 |
Neste caso, as combinações parentais foram 21.379 + 21.906 = 43.285 / 44.595, ou seja, 97,06% do total, enquanto que as recombinações foram 638 + 672 = 1.310 / 44.595, ou seja, 2,94% do total. As proporções entre as combinações parentais e as recombinações foram só ligeiramente inferiores às encontradas no primeiro experimento.
Recombinação ou permutação
Em 1909, o citologista F. A. Janssens (1863-1924) descreveu o fenômeno cromossômico conhecido como permutação ou crossing over, que ocorre durante a prófase I da meiose e consiste na troca de fragmentos entre cromossomos homólogos. Em 1911, Morgan usou as observações de Janssens para explicar a ligação incompleta entre os genes.
Pela teoria de T.H. Morgan, o linkage se deve ao fato de os genes ligados se encontrarem no mesmo cromossomo. Contudo, se os cromossomos permanecessem intatos na herança, os genes situados no mesmo cromossomo deveriam permanecer sempre juntos; em outras palavras, a ligação deveria ser completa. Isso não é realmente o que ocorre, visto que os genes ligados algumas vezes se separam.
Assim, os genes para a cor das sementes, C, e para o endosperma ser liso ou contraído, S, no milho, permanecem ligados nas combinações parentais aproximadamente em 97% dos gametas, enquanto se separam aproximadamente em 3%, conforme visto na tabela 2. Morgan atribuiu a recombinação dos genes ligados à troca de partes entre os cromossomos homólogos (crossing over), à qual chamou de recombinação.
Grupos de linkage e cromossomos
Se um certo gene, A, apresenta linkage com outros dois genes B e C, estes dois últimos também estarão vinculados. Em animais e vegetais podem ser preparados cruzamentos para averiguar se os genes são herdados independentemente ou se há linkage entre pares ou grupos de genes.
Estes experimentos são particularmente simples nas espécies de Drosophila, pois devido à ausência de recombinação nos machos híbridos se verifica imediatamente quais genes estão ligados e quais segregam independentemente. Desta forma, em cada espécie os genes conhecidos podem ser divididos em grupos de linkage. Os membros de cada par de homólogos apresentam os genes em linkage entre si. Nas espécies geneticamente bem estudadas, o número de grupos de ligação é igual ao número de pares de cromossomos, isto é, é igual ao número haploide de cromossomos que a espécie apresenta.
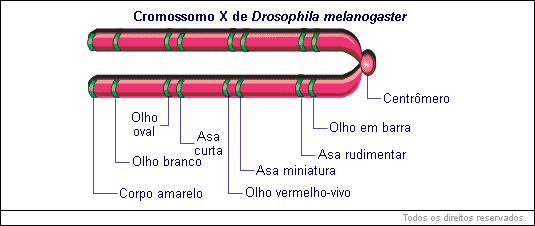
Em Drosophila melanogaster foram observadas muitas mutações desde 1909, desde que esta espécie foi utilizada pela primeira vez nos estudos de T.H. Morgan e seus colaboradores. Os estudos sobre a herança destes tipos mutantes demonstraram que pertencem a quatro grupos de linkage. Lembremos que nestas moscas 2n = 8 cromossomos e que, portanto, n = 4 cromossomos.

Fatores que afetam a força do linkage
A frequência de recombinação (crossing over) entre genes e, por conseguinte, a frequência de formação de quiasmas entre seus loci, no cromossomo, pode ser influenciada por fatores ambientais e fisiológicos.
Assim, ao aumentar a idade das fêmeas de Drosophila melanogaster, diminui a frequência de recombinações na formação dos gametas. A temperatura, os raios X e a composição química do alimento são outros agentes que modificam a frequência das recombinações.
Estimativa da distância entre genes
A frequência ou taxa de recombinação entre dois genes, representa a soma das frequências em que ocorrem as classes recombinantes:
|
frequência de recombinação = soma das frequências das classes recombinantes |
Mapas genéticos dos cromossomos
Em 1923, Alfred Sturtevant, um dos pesquisadores do grupo de Morgan, concluiu que assumindo a probabilidade de ocorrência de permutação como constante ao longo de todo cromossomo, a porcentagem de recombinação entre dois locos do mesmo cromossomo, refletiria a distância entre eles. Portanto, quanto mais distantes estão situados dois genes no cromossomo, maior é a probabilidade da ocorrência de permutação entre eles; se os genes estiverem muito próximos, menor a chance de ocorrer o crossing-over entre eles. Sturtevant concluiu que a porcentagem de recombinação entre dois locos gênicos reflete diretamente a distância entre esses locos no cromossomo.
Uma vez estimada a distância entre dois genes a partir da frequência de permutação, pode ser construído o “mapa gênico” dos cromossomos. Esse mapa representa a sequência dos loci gênicos e suas respectivas distâncias.
Num dos casos estudados por Sturtevant estavam envolvidos três genes mutantes: y, para corpo amarelo, v, para olho vermelho, e m, para asa em miniatura. Já se conhecia a frequência de recombinação entre v e y (32,2%) e entre m e y (35,5%). Portanto, v estava mais próximo de y do que m. Então, qual seria a posição relativa de m e v?
A previsão de Sturtevant era de que a distância tanto poderia ser 67,7% (35,5 + 32,2) como 3,3% (35,5 - 32,2).
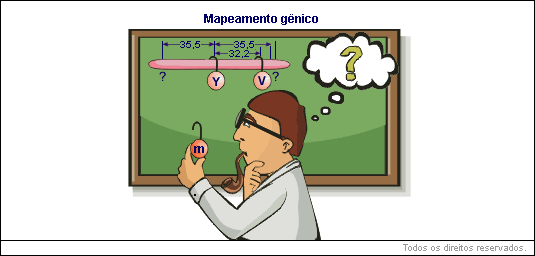
O pesquisador cruzou fêmeas duplo-heterozigotas e machos recessivos para os genes v e m. Obteve uma porcentagem de recombinantes igual a 3%, muito próxima de uma das previsões (3,3%). O resultado ofereceu forte evidência de que as hipóteses consideradas para a construção de mapas genéticos estavam corretas.
A unidade usada como medida de distância entre genes, no mesmo cromossomo, é a unidade de recombinação (UR) ou morganídeo, sendo esta última em homenagem a Morgan.
|
Uma unidade de recombinação, ou morganídeo, corresponde à frequência de recombinação igual a 1%. |
Com esse mesmo procedimento científico, os pesquisadores construíram numerosos outros mapas genéticos com muitas espécies de organismos.
Sumário
- Linkage (ligação ou vinculação)i. Ligação no milho
ii. Recombinação ou Permutação
iii. Grupos de linkage e cromossomos
iv. Fatores que afetam a força do linkage
v. Estimativa da distância entre genes
- Mapas genéticos dos cromossomos
Áreas exclusivas para assinantes