-
HOME
Educação sem fronteiras
-
ALUNOS
A faculdade ao seu alcance

Assine o Educabras!
Próxima parada, faculdade
Nosso objetivo é prepará-lo para o Vestibular e o Enem, para que você possa entrar na faculdade de sua escolha e seguir a carreira de seus sonhos.
Torne-se um assinante a partir de R$29,90/mês Mais Informações

-
PROFESSORES
agilize seu trabalho
Assine o Educabras!
Solução inteligente
Fácil de usar, eficaz e econômico, o Educabras.com é uma solução para professores de Ensino Médio, Pré-Vestibular e Enem.
-
COLÉGIOS
Planos diferenciados

Para colégios, cursos e ONGs.
Colégio conectado
Duas opções:
1. Assinaturas a preços com desconto.
2. Uma única assinatura que inclui o gerador de provas de todas as matérias. -
Área Livre
Aulas e Blog

Área livre do Educabras.com, aberta para assinantes e visitantes.
Educação é essencial
Educação, Conhecimento e diversos outros assuntos relevantes para alunos e professores.
-
Entre em contato
Tire suas dúvidas


Comportamento Social
- Home
- Comportamento Social
Enquanto o organismo individual pode depender de mecanismos adaptativos de instinto, aprendizado e raciocínio para sobreviver, estes quase sempre operam em um contexto social, acima de tudo, a sobrevivência das espécies frequentemente depende de alguma forma de comportamento social.
Muito claramente, uma organização social proporciona seu próprio valor adaptativo para o indivíduo e para a espécie, facilitando a reprodução e a criação dos jovens, fornecendo prevenção e defesa adicional contra predadores e contribuindo para a solução dos problemas de alimento, água, temperatura e outros desafios do ambiente.
Ao mesmo tempo, uma organização social produz seus próprios problemas, contribuindo para tensão emocional e doenças. Em nenhum lugar do reino animal, são os benefícios e as desvantagens de uma organização social tão claramente ilustrados como no caso do homem e suas sociedades. Olhando para a evolução do comportamento social e analisando os mecanismos biológicos e comportamentais que o sustentam, poderemos ser capazes de avaliar os propósitos e limites básicos da sociedade, seus poderes e desorganizações.

Comportamento social, por definição, implica a interação de dois ou mais indivíduos, ou a influência de um indivíduo sobre outro. A simples existência de comportamento de grupo, naturalmente, não garante comportamento social. Por exemplo, a orientação e o movimento de um grupo de paramécios na direção de uma fonte de luz é simplesmente uma agregação de indivíduos respondendo a um estímulo externo comum. Assim são os voos de um grupo de mariposas ao redor de uma lâmpada e a alimentação de moscas em um torrão de açúcar. Cada indivíduo responderia da mesma forma, mesmo que estivesse sozinho.
Quando olhamos para trás no reino animal e examinamos o que está envolvido em comportamento social verdadeiro, mesmo nos casos mais simples encontramos alguns elementos em comum:
1. Até certo grau, interação social é envolvida em comportamento reprodutivo da maioria dos animais e as interações macho-fêmea e genitores-descendentes.
2. Em muitos organismos, a história do desenvolvimento inicial é crítica em preparar o palco para papéis sociais ulteriores, caso isto seja feito através de acontecimentos fisiológicos que determinam a morfologia ( por exemplo, casta de abelha, sexo ), ou de acontecimentos psicológicos que determinam tendência de resposta ( por exemplo, dominância, submissão ).
3. A maioria das espécies tem algum meio de comunicação entre os indivíduos, desde um sinal-estímulo simples, que desperta um padrão instintivo, até um processo simbólico, vocal, como a linguagem.
4. Territorialidade, centralizando-se ao redor do local de reprodução, local do ninho e fonte de alimento, é comum e fornece um ambiente para muitos aspectos do comportamento social.
5. Finalmente, cada espécie tem uma organização social característica, determinando se ela manifestará grupos de acasalamentos permanentes, cuidado prolongado com os jovens; organização familiar, grupal ou de rebanhos; moléstias socialmente induzidas e transmissão cultural de uma geração para outra; em um alto grau, estas características dependem do ritmo de desenvolvimento dos jovens e dos dotes neurológicos e endócrinos das espécies.
Comportamento reprodutivo
É instrutivo olhar para o comportamento reprodutivo, pois, pelo menos neste aspecto, quase todos os animais multicelulares são sociais. Certas espécies de borboletas não mostram comportamento social no seu ciclo de vida até que chegue o tempo de acasalar. Então começa uma corte complicada. O macho para de procurar alimento e espera alerta pela fêmea. Cada vez que uma fêmea passa, o macho voa em perseguição. Se ela pousa e permanece imóvel, ele bate suas asas, abrindo-as e fechando-as, expondo suas manchas. Então faz uma saudação profunda, caminha ao redor da fêmea e coloca seu abdome em contato com os órgãos genitais dela. Depois disto, o comportamento social para; o macho nunca interatua com outras borboletas novamente e a fêmea põe seus ovos em objetos alimentares na grama e abandona-os.
Essas interações – e tantas outras que poderiam ser descritas – entre indivíduos são padrões em grande parte inatos, fortemente dependentes de hormônios, e são despertados por sinais-estímulos altamente específicos fornecidos pelo outro indivíduo. Algumas vezes, o sinal-estímulo é uma transformação morfológica induzida pelos hormônios, como a coloração brilhante do macho do peixe esgana-gata. Outras vezes é uma resposta comportamental, como o saudar de certas borboletas.
A proliferação dos gametas femininos, sem rivalizar com a prodigiosa produção das células masculinas, permanece considerável. O áscaris-fêmea bota 50 milhões de ovos por ano; os peixes marinhos, vários milhões em cada época de postura. Os oceanos estariam repletos de ovos em alguns meses, se todos os ovos de arenque se desenvolvessem. A rã produz de 2.000 a 4.000 ovos por ano. Se todos esses ovos evoluíssem em animais adultos, estes cobririam a terra com uma camada de 40 cm depois de 5 anos.
Também nas aves, o poder vegetativo dos gametas é muito importante. O ovário de uma ave fêmea na eclosão encerra aproximadamente 100.000 ovócitos. O número de ovos postos não ultrapassa 1.000 para as melhores raças selecionadas de galinhas e patas. Essa redução do número de ovos postos depende de duas causas. A fase de multiplicação das ovogônias interrompe-se de 4 dias a uma semana antes da eclosão e, em seguida, no caso das aves selvagens, o período de postura limita-se a uma época bem definida. O peso do ovário de estorninho aumenta de 30 vezes na primavera devido ao crescimento de uma centena de ovócitos. Somente cinco a oito óvulos são formados; os outros ovócitos são absorvidos ao nível do ovário. Entretanto, em certas condições, uma parte importante dos ovócitos grandes pode ser emitida. Se retirarmos diariamente um ovo do ninho da pardoca a partir do segundo dia da postura, esta prosseguirá por muito mais tempo. A fêmea bota até 50 ovos, ao passo que, normalmente, são encontrados não mais que três ou quatro ovos no ninho.
Na espécie humana, os dois ovários da menina ao nascimento encerram várias centenas de milhares de ovócitos, dos quais muitos terão atresia folicular. Entre os que ficam, um pequeno número evoluirá nos folículos terciários desde a puberdade até a menopausa. Durante a vida sexual da mulher, supondo que não haja gravidez nem outros fenômenos inibidores, serão ovulados somente 400 a 500 ovócitos.
De maneira geral, a produção dos ovos se reduz cada vez mais à medida que a proteção do jovem fica mais assegurada e, por conseguinte, suas chances de sobrevivência aumentam. Os peixes marinhos produzem muito mais ovos que a maioria dos peixes de água doce e que os anfíbios; as chances de sobrevivência nos rios, nos lagos ou nas lagoas são bem maiores que no mar. Entretanto nos elasmobrânquios, tais como o tubarão, os ovos se desenvolvem nos ovidutos e o jovem nasce numa fase já bem avançada. Os seis a doze ovos que então evoluem a cada período de reprodução bastam para assegurar a perpetuação da espécie. Os sauropsídeos, cujo jovem desenvolve-se no interior de uma casca protetora mais ou menos calcificada, botam muito menos ovos que os anfíbios. Enfim, os mamíferos que protegem ao máximo os jovens graças ao aparelho placentário só produzem um único ovo em cada ciclo de reprodução.
A sobrevivência dos óvulos é sempre limitada, tratando-se de animais onde a fecundação se realiza no interior das vias genitais femininas ou de animais cuja fecundação é externa. A sobrevivência não ultrapassa alguns segundos ou alguns minutos para os animais aquáticos. É de 3 ou 4 horas no máximo para o ovo de ascídia (urocordado). A mudança estrutural do ovo que pode ser observada em contato com a água impediria rapidamente a fecundação. Não se trata de uma perda de material endógeno do ovócito já que, uma fez fecundado, ele prossegue o desenvolvimento muito antes que o concurso nutritivo externo torne-se indispensável. Por exemplo, o ovo do ouriço-do-mar pode sobreviver duas semanas sem essa ajuda nutritiva. Sua atividade respiratória, entretanto, torna-se dez vezes mais importante que a do ovo não fecundado ( óvulo ). Sua sobrevivência normal é de 36 a 52 horas.
Uma diminuição ligeira do pH ou da porcentagem de cálcio na água permite prolongar o período de fecundação dos ovos de animais marinhos. Os óvulos não fecundados do ouriço-do-mar sobrevivem cinco vezes mais na água do mar estéril. A ação citolítica das bactérias só se manifesta após um certo período de imunidade, pois os ovos colocados num meio rico em microrganismos sobrevivem tanto tempo quanto em condições comuns. Os "ovos" maduros não fecundados de ouriço-do-mar contraem-se numa solução hipertônica. Rugas indicam que um estado semissólido estabelece-se na superfície; posteriormente, ele voltará a ser liso. Os ovos velhos contraem-se, mas a superfície traduz um estado superficial liso.
Parece, então, que é necessária uma certa energia para impedir a destruição da superfície do ovo em período de envelhecimento. O ATP em fraca concentração pode prolongar a sobrevivência e mesmo restaurar ovos velhos. O rápido envelhecimento do óvulo não fecundado é devido à perda ou incapacidade de sintetizar novamente um “transportador’ de energia. Entre elasmobrânquios, répteis e aves onde os espermatozoides devem subir até a região superior dos ovidutos, a fecundação deve-se realizar nos minutos que seguem à ovulação. Entre os salmões e na maioria dos peixes, o óvulo só é fecundável durante os 15 ou 20 minutos que seguem à sua emissão na água. A sobrevivência do óvulo de mamífero não ultrapassa um dia após a ovulação: 2 a 4 horas para a jumenta e a coelha, umas dez horas para a rata, de 4 a 8 horas para o camundongo-fêmea.
A sobrevivência do óvulo humano não deve ultrapassar 24 horas ou até menos, após a postura ovárica. O óvulo da cadela conservaria a sua fecundidade durante 4 ou 5 dias, talvez uma semana.
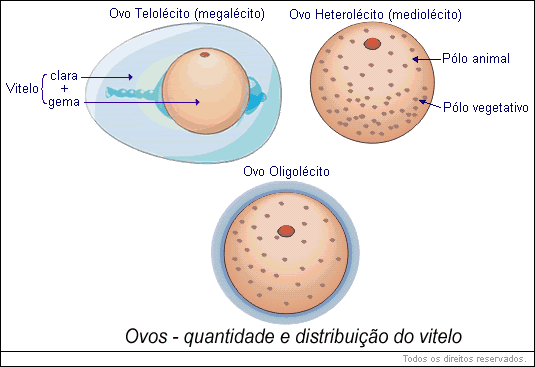
O tamanho dos óvulos não tem nenhuma relação com o tamanho dos animais dos quais derivam; depende apenas da quantidade de vitelo. O diâmetro dos óvulos pobres em substâncias de reserva situa-se nas vizinhanças do décimo de milímetro; os óvulos munidos regularmente de vitelo (mediolécitos), como os dos anfíbios, têm um diâmetro da ordem de milímetro; enfim, para os óvulos muito ricos em vitelo ( megalécitos ), o diâmetro pode atingir um centímetro, às vezes, mais. Assim, o diâmetro da gema do ovo do avestruz mede 7,5 cm. Os ovos dos grandes répteis da era mesozoica deveriam ter alcançado tamanho notável.
Nas aves não só o volume do gameta feminino é muito importante como também as substâncias de reserva acumuladas em função do número de ovos postos, representando um peso notável em relação ao do animal. Assim, uma galinha selecionada da raça Leghorn que pese mais ou menos 2,5 kg pode botar 200 ovos, até mais, durante um ano. Sendo o peso médio de um ovo 55 gramas, a galinha botará mais ou menos 11 kg de ovos, ou seja, quatro vezes seu próprio peso.
Em função dos materiais energéticos acumulados no ovo, o estado de desenvolvimento onde o embrião exige uma carga nutricional vinda do exterior varia conforme os grupos animais. Sendo o ovo de ave o mais rico em substâncias de reserva ( megalécito ), o comportamento geral do ovo na eclosão é o de um adulto imaturo. O ovo de anfíbio (mediolécito) é mais ou menos provido de vitelo. O embrião quando toma a substância nutritiva é ainda uma larva longe da metamorfose. Para o óvulo dos mamíferos placentários, quase desprovido de reservas (oligolécito ), a carga externa deve provir desde as primeiras fases do desenvolvimento, antes de qualquer diferenciação morfológica visível.
Transplante de ovos nos mamíferos
Na prática da inseminação artificial, selecionamos um progenitor macho em razão do interesse econômico de seus caracteres hereditários. Entretanto, há também fêmeas que apresentam qualidades excepcionais, das quais seria interessante obter numerosos descendentes.
Nos bovinos, a produção de grande quantidade de leite por uma vaca, 2 ou 3 vezes superior ao normal, constitui uma grande vantagem na medida em que a aptidão para a lactação comporte-se como um caráter hereditário. Ora, na vaca, um só óvulo é produzido em cada ciclo sexual, que dura 21 dias. Por conseguinte um animal que nunca apresentasse gestação, seria capaz de produzir teoricamente pelo menos 130 a 180 óvulos. Bastaria fecundar esses óvulos desde a sua emissão e transplantá-los para outras vacas que tornariam certo o seu desenvolvimento. Insistimos bastante no ponto de visto totalmente teórico desse exemplo, sem levar em conta os problemas práticos e principalmente fisiológicos apenas suspeitos, que seriam assim levados em conta.
Entretanto, o transplante de óvulos fecundados realiza-se facilmente nos animais de laboratório e em particular na coelha. Podemos prelevar os óvulos fecundados de uma coelha ou até mesmo óvulos virgens que fecundamos in vitro. Esses ovos são em seguida transplantados para o útero de outra coelha em pseudogestação, ou seja, uma coelha anteriormente cruzada com um macho vasectomizado. Do ponto de vista genético, a coelha hospedeira não representa a verdadeira mãe dos coelhos aos quais dará nascimento; ela serve de organismo onde se faz o desenvolvimento de um “ovo estranho”.
O problema biológico mais delicado para se resolver é o de assegurar a conservação eventual de óvulos virgens ou fecundados, muito mais frágeis que os espermatozoides. Esse problema é objeto de pesquisas contínuas em razão de seu incontestável interesse econômico e bioquímico.
História do desenvolvimento
Embora um indivíduo ou uma espécie possam ter uma capacidade inata para um tipo particular de interação social, uma grande parte depende de experiência anterior. De fato, alguns aspectos do comportamento social dependem tão claramente de experiência anterior que eles não aparecerão, ou serão severamente distorcidos, se a experiência faltar. Certos tentilhões precisam ouvir o canto da sua espécie antes que possam cantar o seu padrão completo e assim usá-lo em comunicação adulta normal. Além disso, ele deve ouvir o canto antes da sua primeira estação de reprodução, o que indica haver período crítico para o desenvolvimento do comportamento social.
O mesmo tipo de período crítico em desenvolvimento social verifica-se cedo na vida de muitos organismos. Impressão em aves é um exemplo clássico, no qual em menos de um dia o eclodido pode desenvolver uma forte resposta à mãe ou a uma substituta da mãe. Nos carneiros, o filhote é aceito pela mãe apenas dentro das primeiras 4 horas após o nascimento.
Estudos mais extensos foram feitos com macacos que foram criados com vários tipos de mães artificiais – mães feitas de pelúcia ou mães feitas de arame duro. Comparados com o macaco jovem criado com sua própria mãe, jovens criados com mães substitutas inanimadas artificiais são tímidos na resposta a estranhos ou situações estranhas, não brincam adequadamente com seus companheiros e não se acasalam bem sucedidamente quando se tornam adultos. Se a mãe artificial for feita de arame duro, estas deficiências em comportamento social serão profundas e desabilitantes; se a mãe artificial for feita de pelúcia e enchimento macio, os sintomas serão muito menos marcados. Ainda mais, o jovem desenvolver-se-á melhor se a mãe artificial for feita para se balançar ou se movimentar em vez de permanecer fixa e parada.

Estudos clínicos sugerem que a criança humana pode ser ainda mais dependente de certos aspectos críticos da interação com sua mãe, pois crianças criadas em condições de saúde e alimentares excelentes em orfanatos podem sofrer de falta de afeição. Quando isto acontece, elas fracassam no desenvolvimento e frequentemente sucumbem às moléstias; se sobrevivem, são frequentemente sisudas e socialmente retraídas.

Territorialidade
Observação comum mostra que muitos animais mantêm territórios, os quais eles defendem; mas, apenas depois que foram feitos estudos minuciosos com aves canoras inglesas, no início do século passado, é que foi apreciado quão complexa e quão intensa pode ser a territorialidade. O pardal canoro, por exemplo, assume uma “estação de exibição” em um território, que pode ter o tamanho de um hectare, e aí canta a sua canção com grande persistência. Se outro pardal-macho aparece, ele é expulso agressivamente. À medida que a primavera continua, o cantar e a competição pelo território tornam-se mais intensos e, de fato, o território pode aumentar em mais uma metade do que era, antes que o pareamento com uma fêmea se realize.
Bandos de macacos e chimpanzés, assim como famílias de gorilas, movem-se através de territórios de limite do lar que são claramente seus, embora nem sempre tão bem definidos. Um bando de babuínos de 40 a 80 indivíduos pode se distribuir em uma área de até cerca de 10 quilômetros quadrados e este território pode recobrir os limites de um ou mais bandos vizinhos, especialmente ao redor de uma lagoa. Com 400 babuínos em uma única lagoa, foi registrado que não há qualquer mistura de bandos nem qualquer luta, embora bandos pequenos possam ser subjugados pelos maiores.

Territorialidade, obviamente, desempenha muitas funções. Ela, com certeza, facilita o acasalamento e o cuidado dos jovens, mas também fornece solução habitual dos problemas de alimentação e manutenção de abrigo. A observação mostra também que a territorialidade reduz a agressão ou luta efetiva, exibições ocasionais, necessárias para defesa de um território bem estabelecido e expulsão dos intrusos. Finalmente, como os territórios são necessários para acasalamento e criação dos jovens, é também claro que a territorialidade é um dos fatores que mantêm o controle da população.
Comunicação
Todas as interações descritas nos exemplos de comportamento social aqui oferecidos envolvem comunicação de vários tipos de um animal para outro. Algumas vezes, comunicação é uma exibição, como o “saudar” e o “arrulhar” do pombo-trocaz. Outras vezes, é a redução ou eliminação completa de sinais de exibição agressivos, como a “resposta de rendição” vista em cães, lobos e ratos, por exemplo. Estes animais deitam-se de costas e mostram sua barriga e pescoço quando são derrotados em um encontro agressivo. O atacante não é mais estimulado pelos sinais que despertam o ataque e, assim, para de atacar.
A comunicação não precisa ser sempre visual, auditiva, ou confinada ao presente. Ela pode ser química e, devido a isto, pode, naturalmente, ser antecipada. Assim um cão-macho “marca” seu território para certo tempo no futuro e o intruso encontra um cheiro que tem alguma história. O sagui faz isto não apenas com urina, mas também esfregando glândulas odoríferas localizadas ao redor do órgãos genitais ou no peito. As substâncias químicas envolvidas em comunicação social são chamadas feromônios, por analogia com hormônios; eles são materiais químicos específicos que, em pequenas quantidades, agem sobre outro organismo à distância do seu ponto de liberação e de formas muito específicas.
Naturalmente, a comunicação sonora desperta-nos um interesse especial, pois ela forma a base fundamental da linguagem humana. A linguagem, como sabemos, requer:
1. produção articulada de sons e a capacidade de discriminação auditiva.
2. uma capacidade cerebral básica para processos simbólicos, tanto na produção como na apreciação da linguagem.
3. uma organização social em que a linguagem é usada para comunicação, de acordo com um conjunto de regras que são passadas de uma geração para outra.
A excelente articulação do papagaio e outros pássaros não é linguagem, pois suas vocalizações não são simbólicas e não são usadas como meio primário de comunicação social. Por outro lado, a vocalização dos macacos e outros primatas sub-humanos, embora primitiva quanto às produções de sons, comunica simbolicamente no conjunto social. Por exemplo, é possível reconhecer, pelo menos, de 6 a 12 sons simbólicos diferentes nas interações sociais de macacos, representando diferentes tipos de aviso ou gritos por alimento.
Chimpanzés têm sido treinados a usar gestos simbolicamente nas suas interações com o homem e isto parece ser muito mais fácil e muito mais produtivo que o trabalhoso processo para ensinar palavras – e com sucesso limitado, quase inexistente. É possível que o chimpanzé e outros primatas sub-humanos simplesmente não tenham a capacidade cerebral para produzir sons sob comando e relacioná-los com o significado simbólico apropriado. Neste sentido, eles seriam como uma pessoa que sofreu lesões nas áreas da linguagem do seu cérebro, sendo assim afásicas, não podendo produzir linguagem.
A dádiva da linguagem sozinha coloca o homem à parte do resto dos animais e lhe dá grandes vantagens na adaptação ao seu ambiente e em sua organização social. A linguagem é um instrumento simbólico ilimitado da sua inteligência; no comportamento social do homem, ela é um mecanismo para comunicação e para transmissão cultural de uma geração para a seguinte.
Organização social
Na base de padrões inatos, organizações sociais notáveis têm se desenvolvido, talvez atingindo seu ponto mais alto nos insetos. Formigas têm organizações sociais enormemente complexas, com classes especializadas de indivíduos, com cultivo de “jardins de fungos”, “amamentação” de afídeos e exclusão de estranhos à colônia. Nos vertebrados mais simples – por exemplo, peixes e aves –, certa organização social pode ser vista na migração, nos cardumes, nos bandos e nos rebanhos. Algumas aves mostram uma ordem de bicar, na qual a ave dominante bica a ave mais submissa. Esta ordem de bicar fornece uma organização altamente particular e complicada a um grupo de aves, determina quais indivíduos acasalarão e também satisfaz a função de manter a agressão e o dano físico a um mínimo, uma vez que a ordem seja estabelecida.
Relações de dominância e submissão semelhantes determinam não apenas o tipo de organização social possível, mas também algumas das consequências daquela organização para a saúde dos indivíduos e o tamanho das populações animais. Em um estudo de camundongos mantidos em uma grande gaiola de laboratório – 12 metros quadrados – com um suprimento de alimento e água completamente adequado, verificou-se que, à medida que a população crescia a partir dos 2 pares originais macho-fêmea, os animais mais velhos, os primeiros chegados, tornaram-se os indivíduos dominantes e tinham pelos lustrosos, eram bem alimentados e responsáveis pela maior parte da reprodução. Além desses, camundongos submissos se amontoavam em grandes números ( até 100 ) nos cantos da gaiola, enquanto outros, também submissos, encontraram pequenas plataformas, nas quais podiam sentar e se isolar do resto do ambiente social.
Pormenores surpreendentes das mudanças patológicas que podem ser causadas pela pressão social são vistos em estudos de laboratório com galinhas alojadas em números diferentes e diferentes estados sexuais. Doenças cardíacas, por exemplo, aumentaram à medida que o tamanho do grupo cresceu além do dobro. Quando havia quatro ou cinco machos para cada fêmea, ou vice-versa, o aumento em patologia da artéria coronária foi muito marcado, embora os machos se mostrassem mais suscetíveis que as fêmeas.
Implicações para o homem e suas sociedades são claras. Organização social e contatos sociais que exercem pressão emocional sobre os indivíduos, podem resultar em moléstia e morte prematura. Muitas provas indicam o papel de fatores sociais semelhantes em muitas das chamadas doenças degenerativas da civilização.
Sumário
- Comportamento reprodutivo
- Transplante de ovos nos mamíferos
- História do desenvolvimento
- Territorialidade
- Comunicação
- Organização social
Áreas exclusivas para assinantes